Делене на клетката - Мейоза
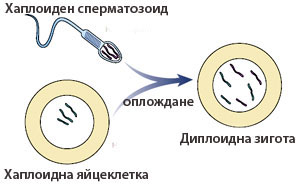 Сложно устроените многоклетъчни организми (цветни растения, животни, хора) водят началото си от една единствена клетка, получена след оплождане, наричана зигота. Тя е резултат от сливането на две хаплоидни полови клетки (яйцеклетка и спермий/ сперматозоид), наричани гамети и носи един набор хромозоми от майчиния организъм и един от бащиния. Формирането на многоклетъчния организъм от една единствена клетка се нарича развитие на организма и се базира на делене и специализация на клетките.
Сложно устроените многоклетъчни организми (цветни растения, животни, хора) водят началото си от една единствена клетка, получена след оплождане, наричана зигота. Тя е резултат от сливането на две хаплоидни полови клетки (яйцеклетка и спермий/ сперматозоид), наричани гамети и носи един набор хромозоми от майчиния организъм и един от бащиния. Формирането на многоклетъчния организъм от една единствена клетка се нарича развитие на организма и се базира на делене и специализация на клетките.
Половото размножаване е една от главните характеристики на еукариотите като група и не се наблюдава при прокариотите. Макар че при някои еукариотни организми отсъства полово размножаване, данните сочат, че те са изгубили тази си способност по пътя на еволюцията.
Половото размножаване включва регулярно редуване на мейоза и оплождане
Мейозата е процес на делене на ядрото, при който хромозомният набор се редуцира от диплоиден (2n) на хаплоиден (n). По време на мейозата, диплоидното ядро на изходната клетка преминава през две последователни деления, първото от които е редукционно. Тези деления резултират в образуването на четири нови клетки, притежаващи намален наполовина хромозомен брой. При растенията мейоза протича при образуването на микроспорите и мегаспорите в цветовете, шишарките или други подобни структури. При животните мейоза протича при образуването на гаметите (яйцеклетки и сперматозоиди).
Оплождането е процес в който две хаплоидни гамети, се сливат и образуват диплоидна зигота. Основна характеристика на половото размножаване е, че новият организъм притежава хромозоми и от двата родителя.
По този начин мейозата контрабалансира ефекта на оплождането, осигурявайки броят на хромозомите да остава постоянен от поколение на поколение.
Размножаване, което включва редуване на мейоза и оплождане се нарича полово размножаване. Жизнените цикли на развитие на всички полово размножаващи се организми включват редуване на диплоидни и хаплоидни хромозомни числа. Различни жизнени цикли се наблюдават в зависимост от времето на протичане на мейозата, след образуване на зиготата.

- При хаплонтните организми (гъби, протисти), зиготата е единствената диплоидна клетка в жизнения цикъл на организма; възрастните организми са хаплоидни. Зиготата се дели чрез мейоза и образува хаплоидни клетки - спори. Тези спори се делят чрез митоза и формират възрастните организми (едноклетъчни или многоклетъчни). Възрастните хаплоидни организми образуват гамети чрез митоза, които се сливат и дават диплоидна зигота.
- При повечето растения се наблюдава редуване на поколенията. При тях мейозта не води да образуване на гамети, а до образуването на спори. Тези спори се делят чрез митоза и формират хаплоидно поколение наречено гаметофит. Върху гаметофита, чрез митоза се формират гаметите. Те се сливат и образуват диплоидна зигота, която се дели чрез митоза и дава диплоидно поколение, наречено спорофит.
- Диплонтните организми (животни и някои растения) са диплоидни и гаметите са единствените хаплоидни клетки в жизнения им цикъл. Гаметите се образуват след мейоза, сливат се и дават началото на зигота.
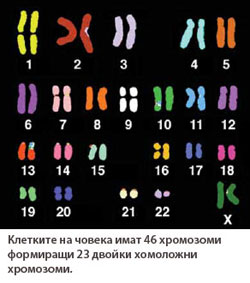
 Всяка диплоидна клетка има по две копия от всяка хромозома, известни като хомоложни хромозоми. Двете хомоложни хромозоми си приличат по размер, форма, мястото на центромерата и вида на генетичната информация която носят, като едната идва от бащината гамета, а втората - от майчината гамета. Например, при човека всяка соматична клетка има 23 двойки хомоложни хромозоми, даващи общия брой от 46 хромозоми.
Всяка диплоидна клетка има по две копия от всяка хромозома, известни като хомоложни хромозоми. Двете хомоложни хромозоми си приличат по размер, форма, мястото на центромерата и вида на генетичната информация която носят, като едната идва от бащината гамета, а втората - от майчината гамета. Например, при човека всяка соматична клетка има 23 двойки хомоложни хромозоми, даващи общия брой от 46 хромозоми.
Мейоза протича само при специализирани диплоидни клетки и то единствено в определен период от жизнения цикъл на съответния организъм. Чрез мейоза и цитокинеза от диплоидната майчината клетка се получават четири хаплоидни клетки - наричани или гамети или мейоспори.
Гамета е клетка, която се слива с друга гамета и дава началото на зигота, а тя от своя страна на нов организъм.
Спора е клетка, която може да даде началото на нов организъм, без да се слива с друга клетка.
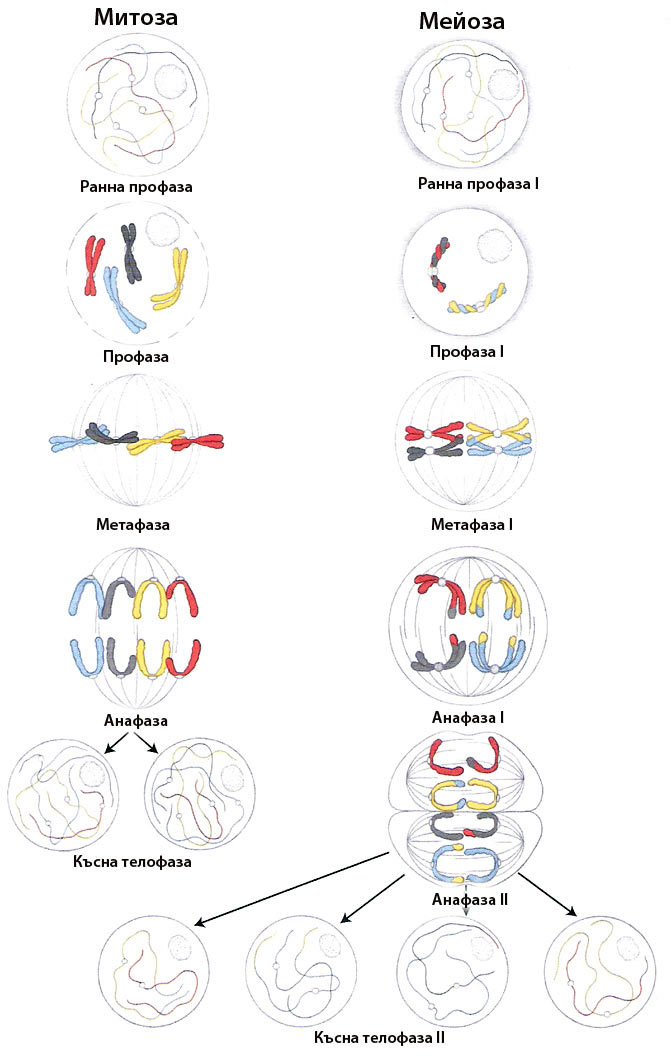
При мейозата ядрото на диплоидната клетка преминава през две последователни деления известни като Мейоза I и Мейоза II, предшествани от само една репликация на ДНК.
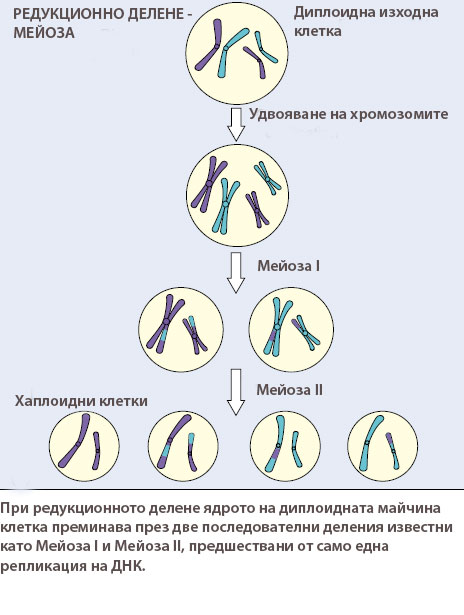 Мейозата е непрекъснат процес, но за по-лесно изучаване се разделя на два етапа, наричани мейоза I и мейоза II. Както при митозата всеки един от тях се подразделя на профаза, метафаза, анафаза и телофаза. При мейозата обаче профаза I е много по-продължителна и сложна.
Мейозата е непрекъснат процес, но за по-лесно изучаване се разделя на два етапа, наричани мейоза I и мейоза II. Както при митозата всеки един от тях се подразделя на профаза, метафаза, анафаза и телофаза. При мейозата обаче профаза I е много по-продължителна и сложна.
Първото мейотично делене редуцира хромозомния брой
Мейоза I започва след приключване на интерфазата. Хромозомите са удвоени и всяка се състои от две идентични сестрински хроматиди, свързани с центромера.
Профаза I
В нея хомоложните хромозоми се асоциират тясно в биваленти посредством синапсис, разменят си сегменти чрез кросинговър, след което се раздалечават оставайки свързани само в отделни точки, наречени хиазми.
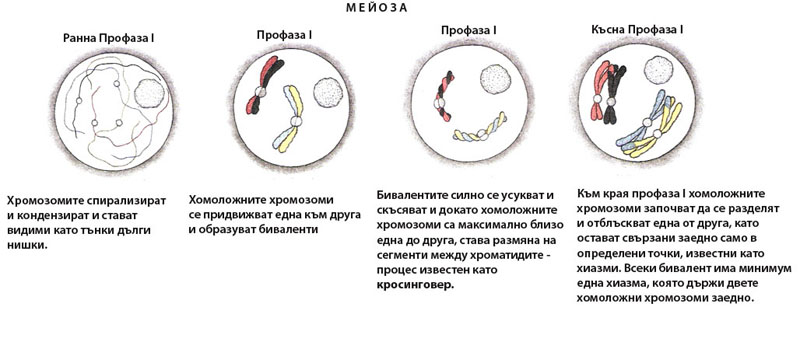
Профаза I започва със спирализиране и кондензиране на хромозомите, които стават видими на светлинен микроскоп като дълги, тънки нишки. Всяка хромозома е удвоена през синтетичния период на интерфазата и се състои от две идентични, свързани заедно, сестрински хроматиди. На този ранен етап на профаза I сестринските хроматиди не са отделени една от друга и удвоените хромозоми изглеждат като единични, а не двойни нишки.
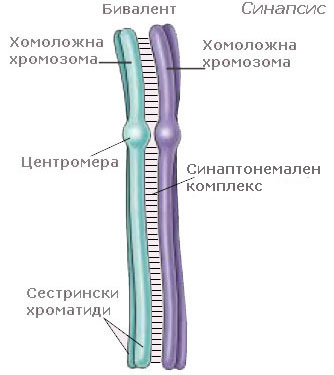
Преди отделните хроматиди да станат видими, хомоложните хромозоми се придвижват една към друга и образуват двойки - известни като биваленти. Формирането на бивалентите е много прецизен процес, при който зоните с еднаква ДНК секвенция в хомоложните хромозоми се разполагат една до друга. Процеса на формиране на двойките хомоложни хромозоми се нарича синапсис и започва от една или няколко точки по дължината на хромозомите и протича по модел на затваряне на цип.
В бивалентите хомоложните хромозоми са разположени плътно една до друга и са устойчиво свързани заедно, посредством субмикроскопична структура наречена синаптонемален комплекс. Той е изграден от протеини и прилича на стълба.
Веднъж формирани, бивалентите продължават силно да се спирализират, кондензират и скъсяват. В този период (когато бивалентите са максимално спирализирани, скъсени и свързани посредством синаптонемалния комплекс) става размяна на сегменти между хроматидите - процес известен като кросинговер. Кросинговерът причинява рекомбинация на наследствените белези и повишава варирането на признаците.
След кросинговера сестринските хроматиди на хомоложните хромозоми вече не са идентични.
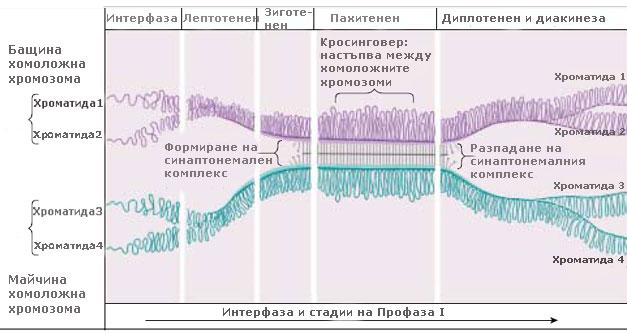
Към края профаза I синаптонемалния комплекс се дезинтегрира и хомоложните хромозоми започват да се разделят и отблъскват една от друга, като остават свързани заедно само в определени точки (местата на рекомбинация), известни като хиазми. Една или повече хиазми могат да се появят върху всяко рамо на хромозомата или може да има само една хиазма за целия бивалент. Задължително обаче ще има поне една хиазма, която да държи двете хомоложни хромозоми заедно.
Профаза I завършва с изчезване на ядърцето и разпадане на ядрената обвивка.
Метафаза I
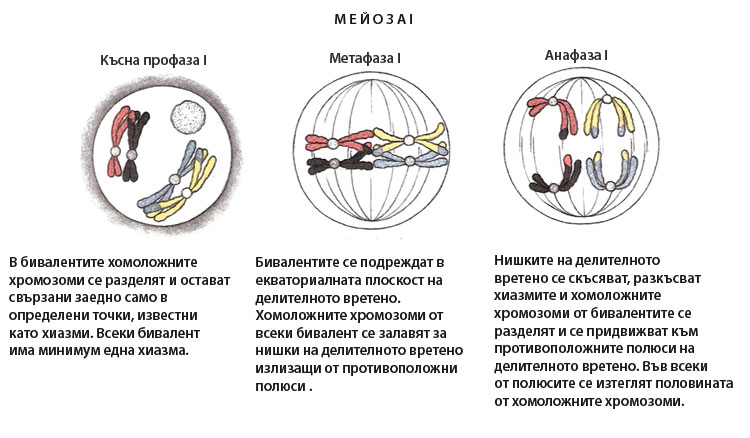
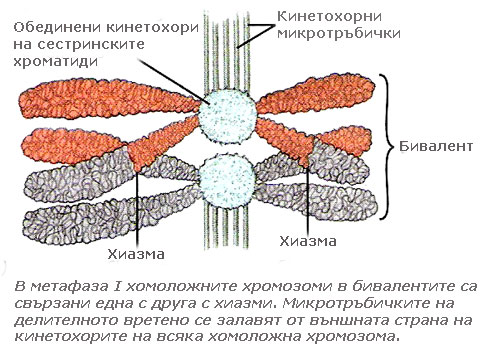 Започва с появата на делителното вретено. Хиазмите държат хомоложните хромозоми заедно, така че само едната страна на центомерата на всяка хомоложна хромозома е обърната навън и достъпна за прикачване към кинетохорните микротръбички. В резултат на това микротръбичките могат да се прикрепят само към външната страна на кинетохорите. Всяка хомоложна хромозома от съответния бивалент, се прикрепя към микротръбички излизащи от противоположни полюси на делителното вретено. Метафаза I завършва с подреждане на бивалентите в екваториалната плоскост на делителното вретено, като е въпрос на шанс коя от хомоложните хромозоми в бивалента към кой полюс на делителното вретено ще се ориентира.
Започва с появата на делителното вретено. Хиазмите държат хомоложните хромозоми заедно, така че само едната страна на центомерата на всяка хомоложна хромозома е обърната навън и достъпна за прикачване към кинетохорните микротръбички. В резултат на това микротръбичките могат да се прикрепят само към външната страна на кинетохорите. Всяка хомоложна хромозома от съответния бивалент, се прикрепя към микротръбички излизащи от противоположни полюси на делителното вретено. Метафаза I завършва с подреждане на бивалентите в екваториалната плоскост на делителното вретено, като е въпрос на шанс коя от хомоложните хромозоми в бивалента към кой полюс на делителното вретено ще се ориентира.
 Анафаза I
Анафаза I
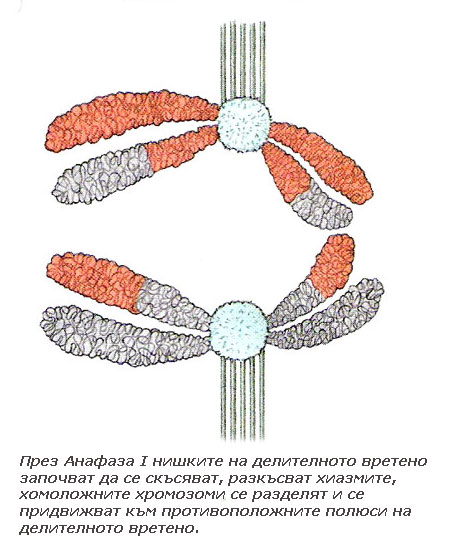
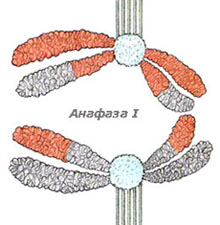
Нишките на делителното вретено започват да се скъсяват, разкъсват хиазмите, хомоложните хромозоми от бивалентите се разделят и започнат да се придвижват към противоположните полюси на делителното вретено. Така във всеки от двата полюса на делителното вретено се изтеглят половината от хомоложните хромозоми. Всяка хомоложна хромозома се състои от две хроматиди, свързани заедно с центромерите си. В края на анафазата хромозомите са разделени наполовина и във всеки от полюсите на делителното вретено има по един пълен хаплоиден хромозомен набор. Понеже хомоложните хромозоми се разполагат на случаен принцип в метафазната пластинка, всеки полюс на делителното вретено може да получи или бащина или майчина хомоложна хромозома.
Телофаза I
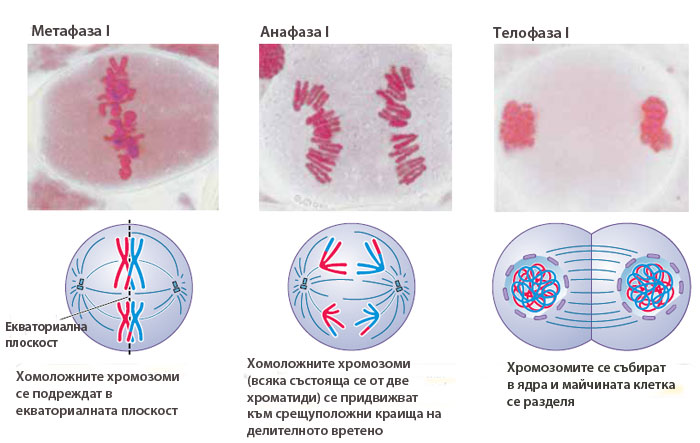 Някои организми имат телофаза I, която протича с образуване на ядрена обвивка. В този случай, след появата на ядрената обвивка хромозомите деспирализират и се удължават, делителното вретено се разрушава, появява се ядърцето и телофазата постепенно преминава в интерфаза, наричана интеркинеза. През нея не се извършва репликация на хромозомите, защото те вече са удвоени и се състоят от две хроматиди.
Някои организми имат телофаза I, която протича с образуване на ядрена обвивка. В този случай, след появата на ядрената обвивка хромозомите деспирализират и се удължават, делителното вретено се разрушава, появява се ядърцето и телофазата постепенно преминава в интерфаза, наричана интеркинеза. През нея не се извършва репликация на хромозомите, защото те вече са удвоени и се състоят от две хроматиди.
При други организми, хромозомите преминават повече или по-малко директно към профаза на второто мейотично делене.
Второто мейотично делене разделя сестринските хроматиди на всяка хромозома
Мейоза II прилича на типично митотично делене, с тази разлика, че не се предшества от репликация ДНК.
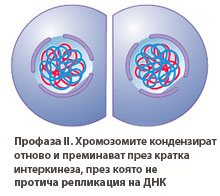 Профаза II
Профаза II
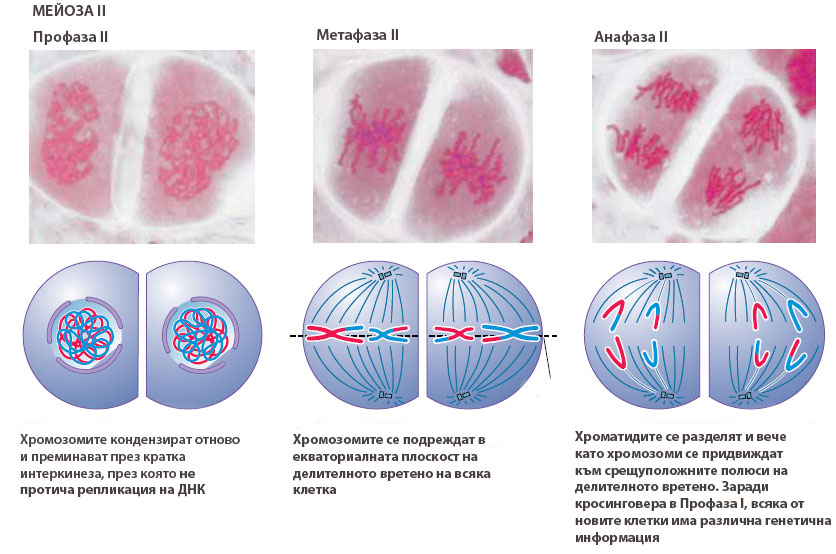
В профаза II хромозомите спирализират отново. В края на профаза II ядрените обвивки на двете ядра се разпадат, изчезват ядърцата (ако са се образували такава през телофаза I) и започва формирането на две делителни вретена. Те могат да бъдат разположени в една плоскост или под различен ъгъл едно спрямо друго
Метафаза II
Нишките делителното вретено се прикрепят към кинетохорите на всяка от двете сестрински хроматиди, така че всяка да е прикачена към срещуположен полюс на делителното вретено. Хромозомите се подреждат в екваториалната плоскост на делителните вретена и формират метафазни пластинки.
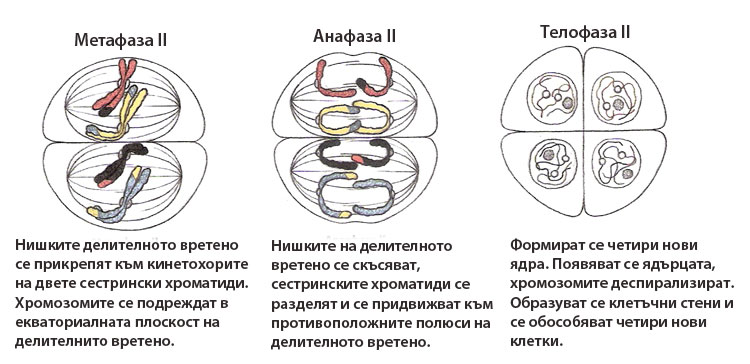
Анафаза II
Нишките на делителното вретено се скъсяват, центромерите се разделят и сестринските хроматиди (наричани вече хромозоми) се придвижват в противоположните полюси на делителното вретено.
Тел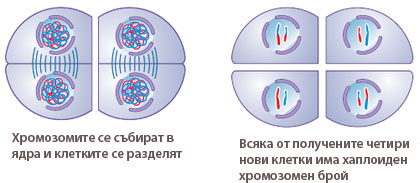 офаза II
офаза II
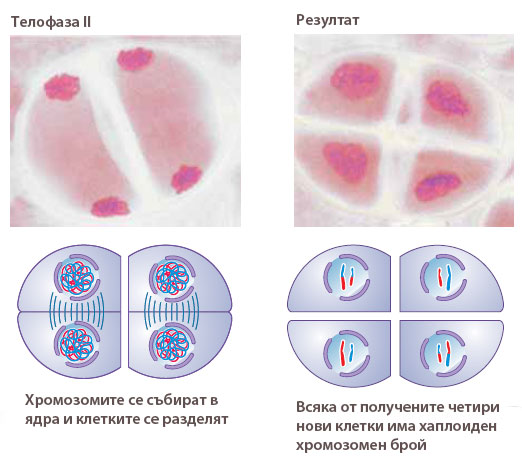
Формират се ядрени обвивки и се обособяват новите четири ядра. Появяват се ядърцата, хромозомите деспирализират и преминават в невидими хроматинови нишки. Образуват се клетъчни стени и се обособяват четири нови клетки.
Мейозата води до генетично разнообразие
Крайният резултат от мейозата са четири клетки с хаплоиден хромозомен набор. Всички те са генетично различни една от друга в резултат на:
- размяната на сегменти между хомоложните хромозоми при кросинговъра, предизвикваща рекомбиниране на гените, и
- произволното ориентиране на бивалентите в метафаза I, така че е въпрос на случайност, коя от хомоложните хромозоми в кой полюс на делителното вретено ще се озове. Например при човека, при наличните 23 двойки хомоложни хромозоми, възможните комбинации при ориентирането им към полюсите са 8 388 608 (формулата е 2 на степен n, в случая 2 на степен 23).
Получените хаплоидни клетки могат да се развият директно в гамети, както е при животните, или в спори, както е при растенията, гъбите и много протисти.
Има три основни разлики между мейозата и митозата
- Независимо че при мейозата протичат две последователни деления на ядрото, те са предшествани само от една репликация на ДНК. Удвояването на ДНК протича преди започване на митозата и мейоза I, но не и преди започване на мейоза II.
- Всяко от четирите ядра получени при мейозата е хаплоидно, т.е съдържа само един хромозомен набор от тези на диплоидната майчина клетка. При митозата двете новополучени ядра имат идентичен диплоиден хромозомен брой като майчината клетка.
- Всяко от четирите ядра получени при мейозата има различна генетична информация от останалите и от изходната клетка, докато ядрата получени чрез митоза имат идентична генетична информация.
Грешки при мейозата
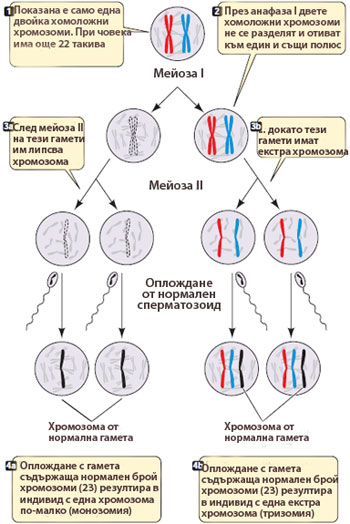 Хомоложните хромозоми могат да не се разделят в мейоза I, или
Хомоложните хромозоми могат да не се разделят в мейоза I, или- Сестринските хроматиди могат да не се разделят в мейоза II
Резултат:
получават се анеуплоидни клетки – клетки, които имат екстра копие на дадена хромозома или недостиг на дадена хромозома.
Анеуплоидията води до генетични увреждания
Например, ако при формирането на човешка яйцеклетка, двете хомоложни хромозоми на хромозома 21 не се разделят и отидат в единия полюс на делителното вретено, две от получените яйцеклетки ще имат по две хромозоми №21, а другите две няма да имат изобщо хромозома 21.
При оплождане на анеуплоидна яйцеклетка от нормален сперматозоид получената зигота:
→ или ще има три копия на хромозома 21 (тризоми) - (ражда се дете с Даун синдром),
→ или ще има само едно копие на хромозома 21 (монозоми)
Тризомите и съответните монозоми са изненадващо чести в човешките зиготи
 10 до 30 % от всички зачевания показват анеуплоидия.
10 до 30 % от всички зачевания показват анеуплоидия.
Повечето от зародишите развиващи се от такива зиготи не оцеляват до раждане, а родените деца от такива зиготи не доживяват до 1 година .- Тризомите и монозомите за повечето хромозоми (с изключение на хромозома 21) са смъртоносни за зародиша .
- 1/5 от всички регистрирани бременности са спонтанно абортирани, заради такива тризоми и монозоми през първите 2 месеца на бременността.
Действителният процент на спонтанно прекъснатите бременности всъщност е значително по-висок, защото ранните бременности остават нерегистрирани изобщо.