Клетка - Клетъчна обвивка
Клетъчна стена
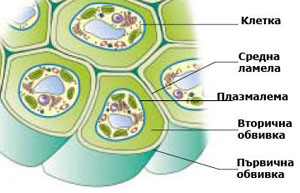 Клетъчната стена е плътна, многослойна структура с полизахаридна природа, която се отлага навън от плазмалемата (плазмената мембрана обграждаща клетката) на растителните клетки. Животинските клетки са ограничени само с плазмена мембрана.
Клетъчната стена е плътна, многослойна структура с полизахаридна природа, която се отлага навън от плазмалемата (плазмената мембрана обграждаща клетката) на растителните клетки. Животинските клетки са ограничени само с плазмена мембрана.
Функция на клетъчната стена
- Определя формата и размера на растителните клетки и допринася за дефиниране на окончателната форма на растителните органи.
- Противодейства на осмотичното налягане и не допуска разкъсването на плазмената мембрана в резултат на прекомерното поемане на вода.
- Определя механичната устойчивост на клетката и съответно и общата устойчивост на растенията – тя е твърдият скелет на клетката.
- Помага на растенията да противостоят на силите на гравитацията.
- Участва в абсорбирането, транспорта и секретирането на вещества.
- Предпазва клетката от инфекции посредством синтез на фитоалексини или синтез и депозиране на вещества (лигнин, кутин и др.), които представляват механична преграда пред гъби и бактерии.
- Свързва съседните клетки заедно.
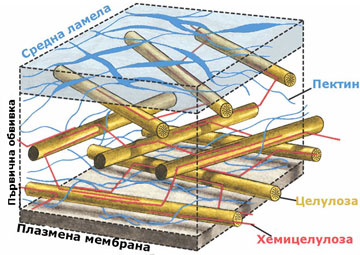
Химичен състав на клетъчната стена
Клетъчната стена е изградена от целулозни микрофибрили потопени в силно хидратиран матрикс от хемицелулози, пектинови вещества и малки количества гликопротеини и липиди.
Целулоза
Целулозата е основният компонент на клетъчната обвивка на растителните клетки и е най-разпространеният структурен полизахарид в растителния свят. 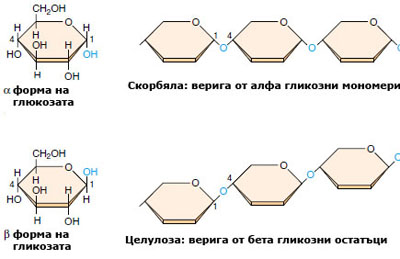 Тя е изградена от свързани помежду си, повтарящи се гликозни мономери. Характеризира се с изключителна механична и химична устойчивост. Неразтворима е във вода, слаби киселини и основи и обикновени органични разтворители. Империчната й формула е идентична с тази на скорбялата - (C6H10O5)n. Във веригата на скорбялата обаче всички гликозни мономери са α-гликоза, докато полимерните вериги на целулозата са изградени от β-гликозни единици. Тази разлика се оказва много съществена, понеже ензимите, които се срещат в повечето организми, лесно разграждат скорбялата, но не са в състояние да разрушават връзката между два β-гликозни мономера. И не защото тази връзка е по-здрава, а защото разрушаването й изисква ензим, който повечето организми не притежават. Изключителната здравина и устойчивост на целулозата я прави много добър биологичен структурен материал.
Тя е изградена от свързани помежду си, повтарящи се гликозни мономери. Характеризира се с изключителна механична и химична устойчивост. Неразтворима е във вода, слаби киселини и основи и обикновени органични разтворители. Империчната й формула е идентична с тази на скорбялата - (C6H10O5)n. Във веригата на скорбялата обаче всички гликозни мономери са α-гликоза, докато полимерните вериги на целулозата са изградени от β-гликозни единици. Тази разлика се оказва много съществена, понеже ензимите, които се срещат в повечето организми, лесно разграждат скорбялата, но не са в състояние да разрушават връзката между два β-гликозни мономера. И не защото тази връзка е по-здрава, а защото разрушаването й изисква ензим, който повечето организми не притежават. Изключителната здравина и устойчивост на целулозата я прави много добър биологичен структурен материал.
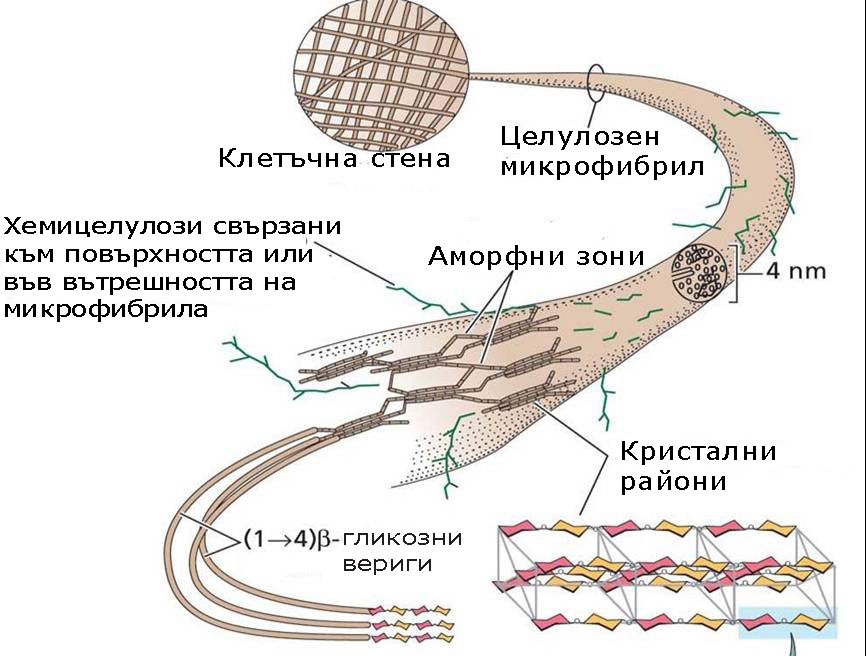 Молекулите на целулозата са дълги линейни вериги в които 300 до 3000 β-гликозни единици се свързват в една целулозна молекула. Тези дълги целулозни молекули имат свойството допълнително да се свързват помежду си чрез водородни връзки.
Молекулите на целулозата са дълги линейни вериги в които 300 до 3000 β-гликозни единици се свързват в една целулозна молекула. Тези дълги целулозни молекули имат свойството допълнително да се свързват помежду си чрез водородни връзки.
- 40-60 целулозни молекули разположени успоредно една на друга образуват елементарни фибрили или мицели, които ясно се виждат на електронен микроскоп.
- 15-20 елементарни фибрили организирани заедно образуват микрофибрили с диаметър от 4-10-25 nm.
- Целулозните микрофибрили от своя страна се усукват заедно, подобно на жичките в кабелите и формират макрофибрили с диаметър 0.5 µm, които лесно се наблюдават със светлинен микроскоп. Усукани по този начин целулозните макрофибрили имат здравина по-висока от тази на стоманени нишки със същата дебелина.
Целулозата има свойства на кристал и подобно на кристалите двойно пречупва светлинните лъчи. Това й свойство се наблюдава в мицелите - зоните където целулозните молекули са подредени успоредно една на друга и имат строга тримерна ориентация. Аморфните зони на микрофибрилите, където гликозни вериги са разположени около и между елементарните фибрили, не проявяват свойства на кристали.
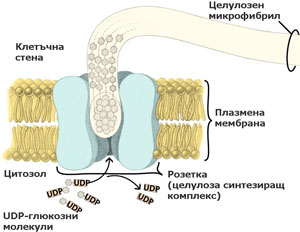 Целулозата се синтезира от целулоза синтезиращи комплекси локализирани в плазмалемата
Целулозата се синтезира от целулоза синтезиращи комплекси локализирани в плазмалемата
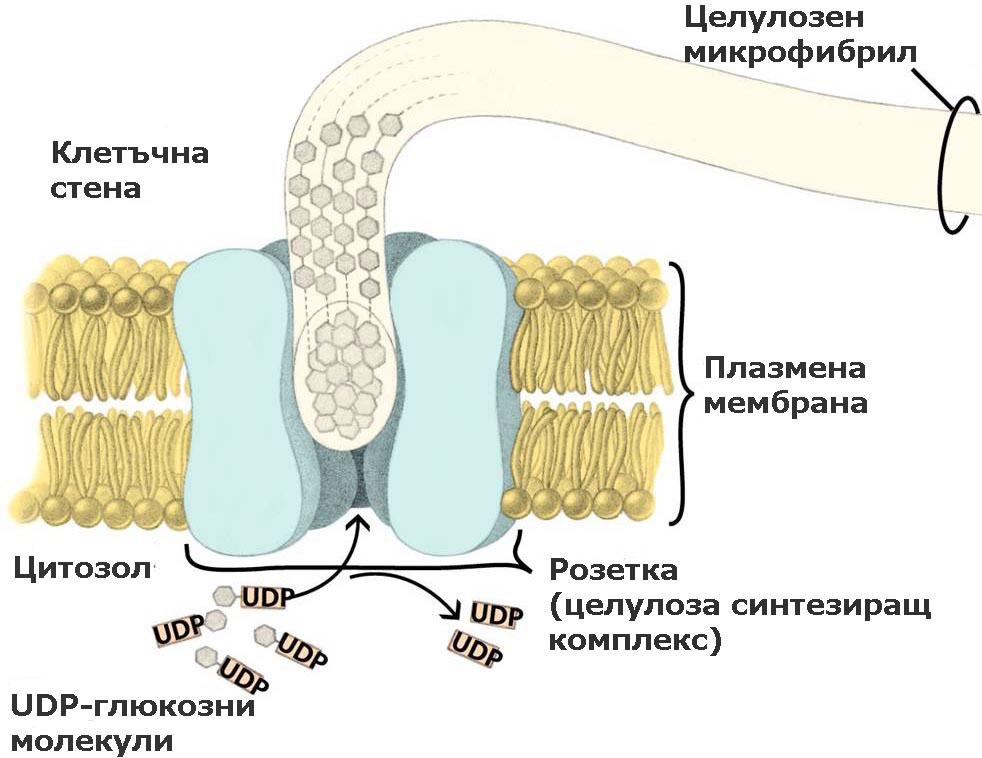
При семенните растения тези ензимни комплекси са организирани като пръстени или розетки от 6 единици, преминаващи през цялата широчина на плазмената мембрана. По време на целулозния синтез розетките се движат през плоскостта на плазмалемата и излъчват образуваните целулозни микрофибрили върху външната повърхност на плазмената мембрана. От там целулозните микрофибрили биват интегрирани в клетъчната стена. Приема се, че движението на розетките се регулира от микротръбички, разположени от вътрешната страна на плазмалемата.
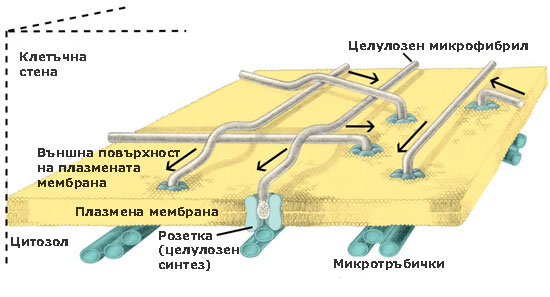 Единиците изграждащи розетките се доставят до и се вграждат в плазмената мембрана (плазмалемата) посредством транспортни мехурчета, образувани от апарата на Голджи.
Единиците изграждащи розетките се доставят до и се вграждат в плазмената мембрана (плазмалемата) посредством транспортни мехурчета, образувани от апарата на Голджи.
Матрикс на клетъчната стена
Матриксът на клетъчната стена е изграден от:
- хемицелулози,
- пектинови вещества и
- гликопротеини
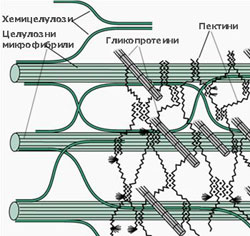 Хемицелулозите са полизахариди с дълги, неразклонени вериги, изградени основно от ксилоза и маноза и малки количества арабиноза и галактоза. Те не формират елементарни фибрили и химичният им състав варира силно, както при различните типове клетки, така и сред различните таксономични групи. Те ограничават еластичността на клетъчната стена, като се свързват посредством водородни връзки с целулозните микрофибрили и фиксират позициите им една спрямо друга.
Хемицелулозите са полизахариди с дълги, неразклонени вериги, изградени основно от ксилоза и маноза и малки количества арабиноза и галактоза. Те не формират елементарни фибрили и химичният им състав варира силно, както при различните типове клетки, така и сред различните таксономични групи. Те ограничават еластичността на клетъчната стена, като се свързват посредством водородни връзки с целулозните микрофибрили и фиксират позициите им една спрямо друга.
Пектините са калциеви и магнезиеви соли на галактуроновата киселина и арабинозата. Те имат силно разклонени молекули, без определена пространствена ориентация. Пектините изграждат изцяло средната ламела, която свързва съседните клетки и се съдържат в големи количества в първичната клетъчна обвивка (до 65%). Те са силно хигроскопични полизахариди и имат способността да поглъщат и задържат значителни количества вода.
Гликопротеини - те са или структурни протеини или ензими. Най-добре познати от структурните гликопротеини са екстензините, които заздравяват клетъчната стена и намаляват разширяването и еластичността й. В първичната клетъчна обвивка е открито и присъствието на голям брой ензими - пероксидази, фосфатази, целулази и пектинази.
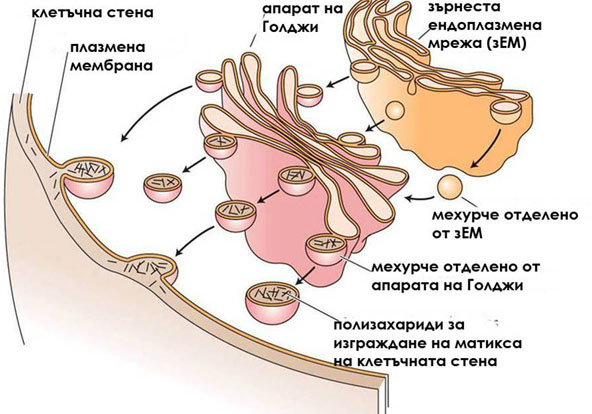
Компонентите на матрикса се синтезират от зърнестата ендоплазмена мрежа и апарата на Голджи
Хемицелулозите, пектините и гликопротеините се синтезират от зърнестата ендоплазмена мрежа и апарата на Голджи, след което се транспортират посредством транспортни мехурчета до плазмалемата. Там те се включват в изграждането на клетъчната обвивка. Типът на синтезираните компоненти на матрикса зависи от етапа на развитие на клетките. Например, пектините преобладават при растящите, удължаващи се клетки, докато хемицелулозите преобладават в клетките които са достигнали окончателните си размери.
Нарастването на клетъчната стена включва взаимодействие между плазмалемата, транспортните мехурчета и микротръбичките
Клетъчната обвивка нараства както на площ така и на дебелина. Нарастването й е комплексен процес, който протича под контрола на протопласта. При нарастването 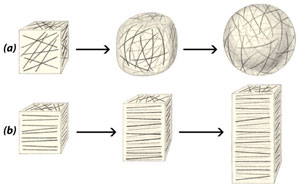 новообразуваните целулозни микрофибрили се отлагат, слой след слой, върху вече образуваните.
новообразуваните целулозни микрофибрили се отлагат, слой след слой, върху вече образуваните.
Начинът на ориентиране на целулозните микрофибрили в първичната клетъчна обвивка повлиява посоката на нарастване на клетките, съответно и формата, която придобиват. Ако целулозните микрофибрили във всички стени на клетката са ориентирани без определен порядък (a), тя придобива сферична форма. Ако целулозните микрофибрили се отлагат перпендикулярно на дългата ос на клетката (b), тя придобива удължена форма.
Много клетки образуват вторична клетъчна стена, в допълнение към първичната
Дебелината на клетъчните стени на растителните клетки варира силно в зависимост от (1) функцията, която те изпълняват и (2) в известна степен и от възрастта на отделните клетки.
Най-рано образуваните целулозни слоеве формират първичната клетъчна стена. Две съседни първични клетъчни стени се свързват със средна ламела (известна още като междуклетъчна субстанция). Много клетки образуват допълнителни слоеве клетъчна обвивка, които формират т.н. вторична клетъчна стена. Ако я има, вторичната клетъчна стена се синтезира и отлага върху повърхността на първичната.
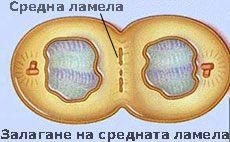 Средната ламела свързва съседните клетки
Средната ламела свързва съседните клетки
При митозата, цитоплазмата на делящата се майчина клетка се разделя с първична преграда, наречена клетъчна пластинка или средна ламела. Средната ламела е изградена основно от пектини и свързва новополучените две дъщерни клетки. Често е трудно да се разграничи средната ламела от първичната клетъчна стена, особено при клетки образуващи дебела вторична клетъчна стена. В тези случаи средната ламела, първичната клетъчна стена и евентуално и първия слой на вторичната клетъчна стена се наричат съставна средна ламела.
Първичната клетъчна стена се отлага докато клетките нарастват
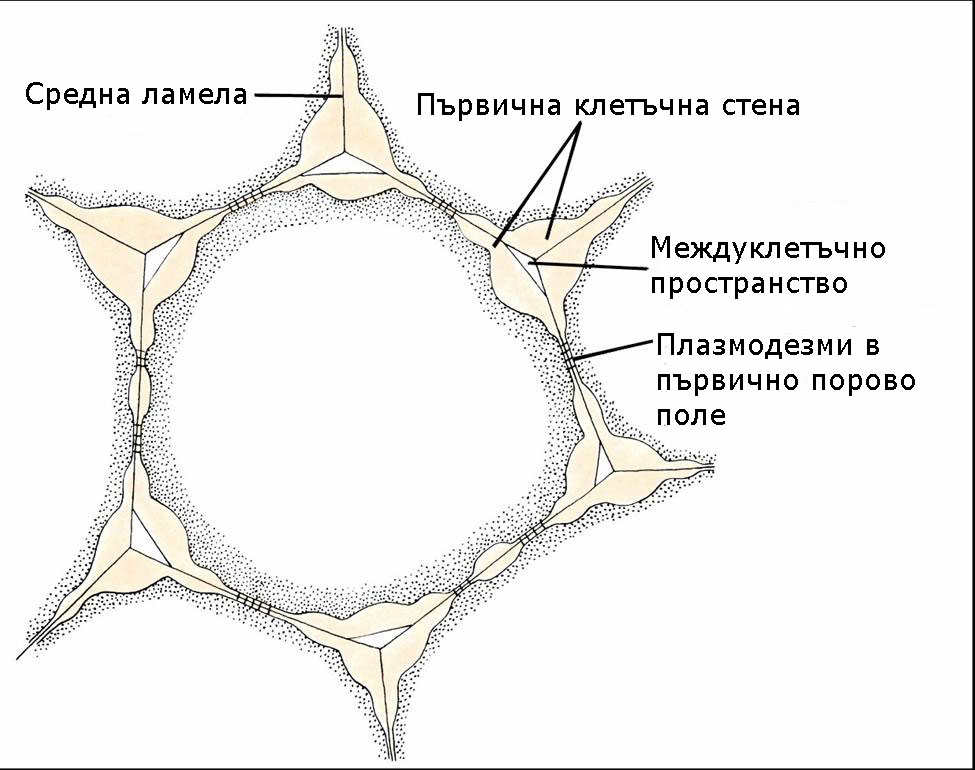 Първичната клетъчна обвивка е характерна за младите растящи растителни клетки. Тя е изградена главно от целулоза, хемицелулози, пектини, протеини (както гликопротеини така и ензими) и вода приблизително в следните съотношения:
Първичната клетъчна обвивка е характерна за младите растящи растителни клетки. Тя е изградена главно от целулоза, хемицелулози, пектини, протеини (както гликопротеини така и ензими) и вода приблизително в следните съотношения:
- целулоза (2-12%)
- хемицелулози (до 20%)
- пектинови вещества
- вода (до 60%)
Бидейки относително тънка и силно еластична, първичната клетъчна стена позволява безпрепятствено повърхностно нарастване на клетката.
Активно делящите се клетки, както и клетките участващи във фотосинтезата, дишането и секретирането на различни вещества, обикновено имат само първична клетъчна обвивка. Последните три вида клетки притежават способността да се дедиференцират и да придобиват способността да се делят отново, след което се диференцират в нов тип клетки. Поради това, принципно само живите клетки с първична клетъчна обвивка участват в заздравяването на раните и регенерирането на тъканите на растенията.
Първичните клетъчни стени също могат да съдържат лигнин, суберин и кутин.
Обикновено първичните клетъчни стени не са с еднаква дебелина по цялата си повърхност. Те имат по-тънки зони наричани порови полета. Плазмодезмите, които свързват протопласта на съседните клетки, обикновено са концентрирани в тези зони.
Вторичната клетъчна стена се отлага след приключване растежа на клетката и на първичната клетъчна стена
Някои клетки имат само първична клетъчна стена, но при други протопластът образува и вторична клетъчна стена. Тя се отлага навътре от първичната клетъчна стена и е локализирана между нея и плазмалемата. Вторичната клетъчна стена обикновено е характерна за специализирани клетки с механична или проводяща функция – коленхима, склеренхима, трахеи, трахеиди. Много от тези клетки умират след отлагането на вторичната клетъчна стена.
Целулозата е основен структурен материал във вторичната клетъчна стена, докато пектините са малко или дори могат да липсват. Това прави вторичната клетъчна стена твърда и неразтеглива. Матриксът й е изграден основно от хемицелулози. Структурните протеини и ензимите, които присъстват във първичната клетъчна стена, липсват във вторичната.
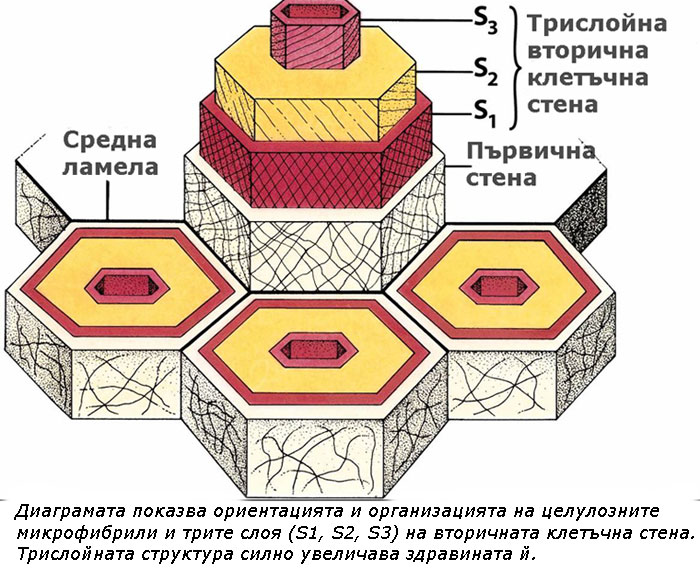 Често три слоя, обозначавани като S1, S2 S3 (за външния, средния и вътрешния слой съответно) могат да бъдат разграничени във вторичната клетъчна стена. Тези слоеве се различават един от друг по ориентацията на целулозните микрофибрили. Трислойната структура на вторичната клетъчна стена силно увеличава нейната здравина. Такива многослойни клетъчни стени се наблюдават при определени клетки на вторичната дървесина и обикновено съдържат и лигнин.
Често три слоя, обозначавани като S1, S2 S3 (за външния, средния и вътрешния слой съответно) могат да бъдат разграничени във вторичната клетъчна стена. Тези слоеве се различават един от друг по ориентацията на целулозните микрофибрили. Трислойната структура на вторичната клетъчна стена силно увеличава нейната здравина. Такива многослойни клетъчни стени се наблюдават при определени клетки на вторичната дървесина и обикновено съдържат и лигнин.
Където първичната клетъчна стена има порови полета, вторичната клетъчна стена има пори
При формирането си вторичната клетъчна стена не се отлага върху зоните, където са разположени поровите полета на първичната клетъчна стена. В резултат на това, характерни прекъсвания, известни като пори, се формират във вторичната стена. В някои случаи пори се формират и в зони където няма първични порови полета.
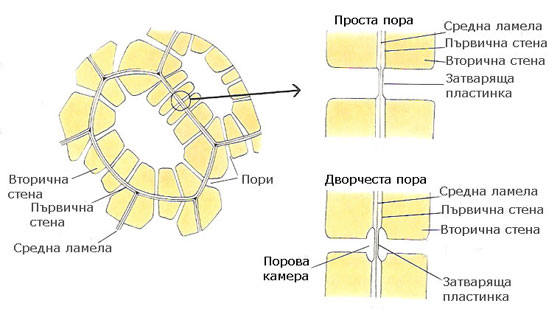
Порите на две съседните клетки обикновено възникват една срещу друга и формират двойки от пори. Средната ламела и двете първични клетъчни стени между две срещуположни пори се наричат затваряща пластинка.
Различаваме два основни типа пори - прости и дворчести
- При простите пори поровият канал е цилиндричен, елипсовиден или цепнатовиден; проекцията му е кръгче или елипса.
- При дворчестите пори се наблюдава надвесване на вторичната клетъчна обвивка над затварящата пластинка и поровият канал е като пресечен конус; проекцията е два вписани кръга.
Лигнифицираните вторични клетъчни стени са непропускливи за вода, но с формирането на порите, съседните клетки са разделени само чрез първичните клетъчни стени в зоната на затварящите им пластинки. Транспортът на веществата се извършва през затварящите пластинки на порите.
Отлагането на вторичната клетъчна стена се нарича още надебеляване на клетъчната стена
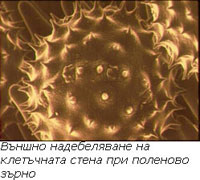
Надебеляването на клетъчната стена бива външно и вътрешно.
Външно надебеляване на клетъчната стена
Наблюдава се при клетки със свободна повърхност (спори, полен) и има формата на мрежи, шипчета, кукички и пр. То е типично за отделните видове и е важен систематичен белег.
Вътрешното надебеляване на клетъчната стена бива скулптурно и поресто
При скулптурното 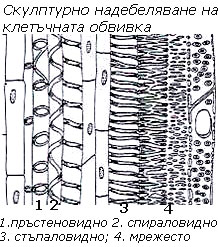 надебеляване надебелените участъци заемат по-малка част от повърхността на клетъчната обвивка и се отлагат под формата на някаква фигура (скулптура). Според формата на скулптурите този вид надебеляване бива:
надебеляване надебелените участъци заемат по-малка част от повърхността на клетъчната обвивка и се отлагат под формата на някаква фигура (скулптура). Според формата на скулптурите този вид надебеляване бива:
- пръстеновидно,
- спираловидно,
- стъпаловидно и
- мрежесто
При порестото надебеляване на клетъчната стена надебелените места заемат по-голяма част от обвивката и само малки участъци, наричани пори, остават ненадебелени. Транспортът на веществата се извършва през порите.
Плазмодезмите са цитоплазмени нишки, които свързват протопласта на съседните клетки
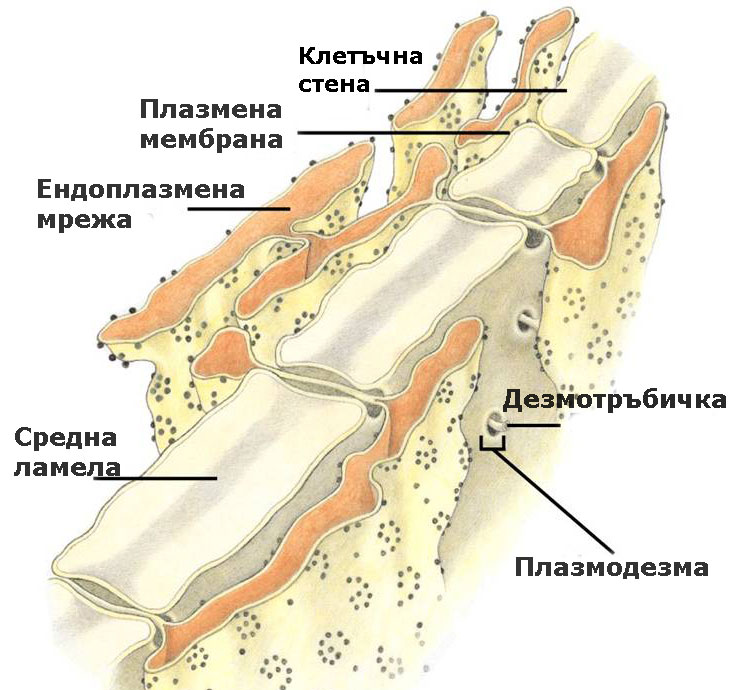 Цитоплазмата на съседните клетки е свързана посредством плазмодезми, които обикновено преминават през първичните порови полета или порите на клетъчните стени и участват в транспорта на веществата и в предаването на сигнали между клетките.
Цитоплазмата на съседните клетки е свързана посредством плазмодезми, които обикновено преминават през първичните порови полета или порите на клетъчните стени и участват в транспорта на веществата и в предаването на сигнали между клетките.
Под електронен микроскоп, те се наблюдават като тесни каналчета с диаметър от 30-60 nm.
Плазмодезмите са тесни каналчета, покрити с плазмена мембрана и изпълнени с цитозол и разположена в средата дезмотръбичка.
Дезмотръбичката представлява модифицирана, тясна и плътна тръбичка на ЕМ, без лумен. Целия транспорт на веществата става през обграждащия я цитозол.
Понеже клетките са свързани посредством плазмодезмите, протопластът на цялото растение, представлява единно цяло и се нарича симпласт. Съответно, движението на веществата през плазмодезмите, се нарича транспорт на веществата по симпласта.
От друга страна, клетъчните стени също образуват единно цяло, наричано апопласт. Движението на веществата по клетъчните стени, се нарича транспорт на веществата по апопласта.
Повечето плазмодезми се формират по време на делене на клетката, като нишки от 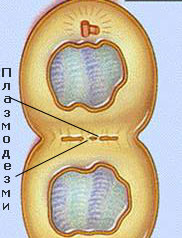 ендоплазмената мрежа, се оказват оградени от залагащата се средна ламела. Плазмодезми могат да се залагат и de novo във вече формираните клетъчни стени. Плазмодезмите формирани през цитокинезата се наричат първични плазмодезми, а тези формирани след цитокинезата - вторични плазмодезми.
ендоплазмената мрежа, се оказват оградени от залагащата се средна ламела. Плазмодезми могат да се залагат и de novo във вече формираните клетъчни стени. Плазмодезмите формирани през цитокинезата се наричат първични плазмодезми, а тези формирани след цитокинезата - вторични плазмодезми.
Броят на плазмодезмите варира и може да достигне до 20 000 в клетка.
Изменения в химичния състав на клетъчните стени
В процеса на развитие на клетката, клетъчната й стена се изменя съществено. Тези промени могат да се дължат на промени в основните компоненти, които изграждат клетъчната обвивка или на допълнително отлагане на нови химични съединения. Новите вещества могат да се отлагат в матрикса на клетъчната стена между целулозните микрофибрили (процеса се нарича инкрустиране) или да се отлагат върху целулозните микрофибрили (процеса се нарича адкрустиране).
Лигнифициране или вдървесиняване на клетъчната стена
 Изразява се в отлагането на лигнини в клетъчните стени. Лигнините са фенолни полимери, изключително широко разпространени в природата - втори след целулозата. Съотношението на мономерите, които ги изграждат (p-кумарил, кониферил и синапил алкохоли), варира значително, както при различните голосеменни и покритосеменни растения, така и при отделните растителни видове, органи и тъкани. Лигнините имат следната роля:
Изразява се в отлагането на лигнини в клетъчните стени. Лигнините са фенолни полимери, изключително широко разпространени в природата - втори след целулозата. Съотношението на мономерите, които ги изграждат (p-кумарил, кониферил и синапил алкохоли), варира значително, както при различните голосеменни и покритосеменни растения, така и при отделните растителни видове, органи и тъкани. Лигнините имат следната роля:
- Придават твърдост и здравина на клетъчните стени. Смята се, че са изиграли основна роля в еволюцията на сухоземните растения. Отлагането на лигнини е дало необходимата механична устойчивост на растенията да противодействат на силите на гравитацията и да развият изправени и силно разклонени стъбла, способни да поддържат големи фотосинтетични повърхности.
- Правят клетъчните стени водонепропускливи и подпомагат възходящия транспорт на водата в проводящите елементи на ксилема.
- Защитават растенията. При нападения от гъби и бактерии или при наранявания се наблюдава отлагане на лигнин в съответните зони. Така наречения раневи лигнин предпазва растението, като повишава здравината на клетъчните стени и те противодействат, както на физическото проникване на гъбите в растителните тъкани, така и на дифузията на ензимите и токсините отделяни от тях. Някои учени смятат, че при еволюцията на сухоземните растения, лигнините първоначално са действали като антибактериални и антигъбни агенти и едва по-късно са придобили съществена роля в транспорта на веществата и повишаване на механичната устойчивост на растенията.
Лигнифицирането на клетъчните стени обикновено води до загиване на протопласта и се наблюдава основно при проводящите и механичните тъкани на растенията.
 Вкорковяване или суберинизиране на клетъчната стена
Вкорковяване или суберинизиране на клетъчната стена
Представлява адкрустиране на суберин в клетъчната обвивка. Суберинът е липидно съединение отличаващо се с голяма химична устойчивост. Вкорковените клетъчни стени не пропускат вода и газове и протопласта загива. Суберинът е главен компонент на стените на корковите клетки, изграждащи най-външния слой на кората на растенията. На електронен микроскоп в суберинизираните клетъчни стени се наблюдават редуващи се светли и тъмни слоеве. Смята се, че светлите слоеве са изградени от восъци, а тъмните - от суберин.
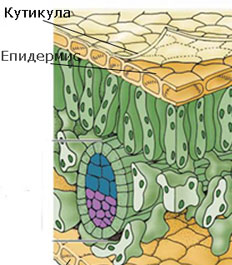 Кутинизация на клетъчната стена
Кутинизация на клетъчната стена
Кутинът е липидно вещество, което е важен структурен компонент на клетъчните стени на много растения. Основната му функция е да формира матрикс, в който се разполагат восъци (липидни съединения с високо молекулно техло и дълги вериги). Восъците в комбинация с кутин формират защитен слой върху външните стени на епидермалните клетки, известен като кутикула. Освен восъци вградени в кутина (кутикулярни восъци), кутикулата често е покрита и от повърхностен слой от восъци (епикутикулярни восъци). Кутикулата (1) предотвратява загубата на вода и други молекули от повърхността на растителните органи и (2) предпазва растенията и от бактериални и гъбни заболявания, като не позволява задържането на вода върху растителните органи.
Мацерация
Представлява разединяване на клетките, вследствие на разрушаване на свързващата ги средна ламела. Бива естествена и изкуствена. Естествената мацерация протича под въздействието на ензима пектиназа, който разгражда пектиновите вещества на средната ламела. Причинява окапването на листата, венчелистчетата, узрелите плодове, както и отделянето на техническите влакна на лена и конопа. Изкуствена мацерация може да бъде предизвикана чрез въздействие с висока температура, синтетични ензими, разредени киселини. При варенето на кореноплоди, плодове, картофи и други продукти от растителен произход винаги настъпва мацерация, в резултат на което те омекват.
 Ослизяване на клетъчната стена
Ослизяване на клетъчната стена
Слизестите вещества представляват хетерополизахариди със способност силно да набъбват във вода. Те се синтезират от растенията с цел поглъщане и задържане на вода. Получават се чрез трансформиране на целулоза, скорбяла и пектинови вещества и се локализират в определени клетки в растителните тъкани. Понякога слизестите вещества се отлагат направо върху клетъчната обвивка като вторично надебеляване – явление което се наблюдава при семена на лен и дюля. Слизестите вещества подобряват контакта на семената с почвените частици, а също поглъщат и задържат на вода, с което улесняват покълването на семената.
Гумозис на клетъчната стена
 Гумите са хетерополизахариди и обикновено се образуват в резултат на нараняване на кората на дърветата. Процесът е известен като “гумозиc” и при него клетъчните стени, цели клетки, а понякога и цели групи от клетки в зоната на нараненото място се втечняват и формират вискозен, лепкав, колоиден разтвор. Този колоиден разтвор формира защитен покривен слой върху наранената тъкан и не позволява проникване на гъби и бактерии през нараненото място. Впоследствие гумозните вещества изсъхват и се втвърдяват по повърхността на кората като различни по форма и цвят полупрозрачни зърна или късчета. Гумозис се наблюдава при слива, джанка, череша, бадем и др. При някои растения, синтезът на гумите може да стане и без патологична причина, като част от механизъм за задържане на вода. Наблюдава се при растения развиващи се в сухи местообитания и освен в кората може да протича и в други части на растението - корени, семена, плодове.
Гумите са хетерополизахариди и обикновено се образуват в резултат на нараняване на кората на дърветата. Процесът е известен като “гумозиc” и при него клетъчните стени, цели клетки, а понякога и цели групи от клетки в зоната на нараненото място се втечняват и формират вискозен, лепкав, колоиден разтвор. Този колоиден разтвор формира защитен покривен слой върху наранената тъкан и не позволява проникване на гъби и бактерии през нараненото място. Впоследствие гумозните вещества изсъхват и се втвърдяват по повърхността на кората като различни по форма и цвят полупрозрачни зърна или късчета. Гумозис се наблюдава при слива, джанка, череша, бадем и др. При някои растения, синтезът на гумите може да стане и без патологична причина, като част от механизъм за задържане на вода. Наблюдава се при растения развиващи се в сухи местообитания и освен в кората може да протича и в други части на растението - корени, семена, плодове.
 Минерализиране на клетъчната стена
Минерализиране на клетъчната стена
Представлява инкрустиране на различни минерални вещества в клетъчните стени. Най-често това са силициев двуокис и калциев карбонат. Минерализирането на клетъчните стени прави растителните органи твърди, грапави и намалява фуражната им стойност. Има защитна функция за растенията, като ги предпазва от неприятели и тревопасни животни. Наблюдава се при хвощовете, житните растения, видовете от сем. Boraginaceae (Грапаволистни) и сем. Brassicaceae (Зелеви) и др.